Pseudogener -er ikke pseudo lengre
 En av de tidligere bevis for biologisk evolusjon i menneskelig genom har vært konseptet om nedarvede 'delte feiltakelser' med sjimpanser, relatert til 'pseudogener' eller søppel-DNA. En trodde tidligere at disse DNA-sekvensene var fossilt avfall i vårt genom, med sine alderdommelige gjenlevninger. Lik mange andre evolusjonære fortolkninger, er også dette blitt snudd opp ned av nye oppdagelser om gen-funksjonalitet (1).
En av de tidligere bevis for biologisk evolusjon i menneskelig genom har vært konseptet om nedarvede 'delte feiltakelser' med sjimpanser, relatert til 'pseudogener' eller søppel-DNA. En trodde tidligere at disse DNA-sekvensene var fossilt avfall i vårt genom, med sine alderdommelige gjenlevninger. Lik mange andre evolusjonære fortolkninger, er også dette blitt snudd opp ned av nye oppdagelser om gen-funksjonalitet (1).
Og selvsagt er det også tilfeller av reelle brutte lenker, knyttet til degenerasjon, men disse sekvensene innehar ikke merkene etter felles arv, som det ofte hevdes. I stedet forsvarer det påstander om at genomet er i en tilstand av informasjons-ødeleggelse (genetisk entropi), ikke progressiv evolusjon.
Bilde 1. Pseudogener produserer regulerende RNA-molekyler
Pseudogener kommer i atskillige former avhengig av fravær/nærvær av visse DNA-sekvens karakteristika. en kategori kalles 'ikke-prosessert pseudogen', som har alle vanlige deler til et protein-kodende gen, men som opprinnelig ble tenkt funksjonsudyktig, på grunn av formodede kodings feil i DNA -noe som forvrenger produksjonen av funksjonelt protein. Men det oppdages nå at pseudogener produserer regulerende RNA-molekyler, som er nøkkel-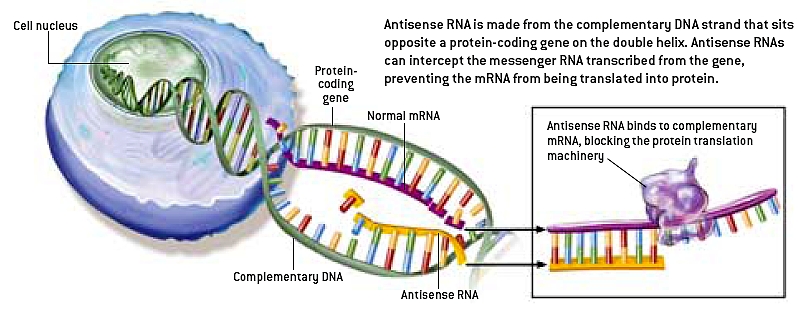 elementer i å kontrollere virkemåten til andre gener og gen-nettverk. Den evolusjonære hypotesen har vært at pseudogener er oppstått hovedsakelig som følge av å ha blitt duplisert fra et annet gen.
elementer i å kontrollere virkemåten til andre gener og gen-nettverk. Den evolusjonære hypotesen har vært at pseudogener er oppstått hovedsakelig som følge av å ha blitt duplisert fra et annet gen.
Bilde 2. 'Antisense' RNA.
Beta-globin pseudogenet -ett case-studie
Beta-globin pseudogenet (HBBP1) er involvert i daglig vedlikehold og regulering av hemoglobin. Dette har vært brukt av evolusjonister som et klassisk argument for felles avstamning mellom mennesket og sjimpanser. Hemoglobinet som finnes i røde blodceller transporterer oksygen rundt i kroppen gjennom sirkulasjonssystemene (store og lille blodomløp). Menneskelig hemoglobin består av en klynge av to ulike kjeder av forskjellige proteiner.
 En av disse kjedene er kalt alpha-globin, som beholdes likt gjennom forløpet fra embryo til voksen 82). Det andre settet kalles beta-kjeden, som spesifikt endrer seg i sammensetning gjennom overganger fra embryo til voksen. HBBP1 pseudogenet er inni en klynge av 6 gener, i menneskelig kromosom 11, som består av over 80.000 baser (2). Se fig. 1. Mens de andre 5 genene i Beta-globin klyngen koder for ett protein, gjelder ikke det HBBP1. Selv om evolusjonister opprinnelig hevdet at genet inneholdt delte kode-feil, så visste de også korrekt nok at genet ville vært fri til å mutere i løpet av de formodentlig 2-6 mill. år av menneskelig evolusjon. I standard evolusjonsmodeller, så er gener som er høyst like sagt å være meget bevarte å være en sterk indikator på funksjonell virkemåte, ellers ville de mutere fritt, uten noe selektivt press på seg. Fra et skapelses-ståsted vil også mene at høy gensekvens-likhet indikerer funksjonalitet, men at det stammer fra gjenbruk av fungerende komponenter for et felles designet formål.
En av disse kjedene er kalt alpha-globin, som beholdes likt gjennom forløpet fra embryo til voksen 82). Det andre settet kalles beta-kjeden, som spesifikt endrer seg i sammensetning gjennom overganger fra embryo til voksen. HBBP1 pseudogenet er inni en klynge av 6 gener, i menneskelig kromosom 11, som består av over 80.000 baser (2). Se fig. 1. Mens de andre 5 genene i Beta-globin klyngen koder for ett protein, gjelder ikke det HBBP1. Selv om evolusjonister opprinnelig hevdet at genet inneholdt delte kode-feil, så visste de også korrekt nok at genet ville vært fri til å mutere i løpet av de formodentlig 2-6 mill. år av menneskelig evolusjon. I standard evolusjonsmodeller, så er gener som er høyst like sagt å være meget bevarte å være en sterk indikator på funksjonell virkemåte, ellers ville de mutere fritt, uten noe selektivt press på seg. Fra et skapelses-ståsted vil også mene at høy gensekvens-likhet indikerer funksjonalitet, men at det stammer fra gjenbruk av fungerende komponenter for et felles designet formål.
Funksjonalitet for HBBP1 har klart blitt påvist. Nyere forskning bekrefter at HBBP1-regionen til 6-geners-Beta-globulin klynger er tydelig ikke-variabelt, sammenlignet med andre beta-globin gener (4). Punktmutasjoner i menneskelig HBBP1 er knyttet til arvelige blodsykdommer (2). Direkte biokjemisk bevis for funksjonalitet for HBBP1 er støttet av multiple kategorier av funksjonelle genomdata, som viser at det er aktivt i å skrive av(kopiere) og en nøkkel-komponent i Beta-globin nettverket (5). Kort sagt koder HBBP1 for to samsvarende RNA-regulatorer, som enten skrives av (kopieres) og/eller spleises på ulike måter. Hele 58 avskriftsvarianter dannes ut fra dette. HBBP1 genet har også fleste forbind-elser med åpne og aktive DNA-regioner innen hele Beta-globin klyngen. 'Pseudogenet' kan med andre ord være det mest aktive i hele klynga av beta-globin gener.
Forbløffende nok uttrykkes HBBP1 gen-avskrifter i minst 250 ulike menneske celle- og vevs-typer. I stedet for å være et ubrukelig genom-fosssil, i følge evolusjonære antagelser, så synes HBBP1 genet å være høyst funksjonelt og smart integrert i det menneskelige genom, men intolerant overfor mutasjoner. Se (2) for mer detaljerte beskrivelser.
 'Retrogener' er også funksjonelle
'Retrogener' er også funksjonelle
En annen kategori pseudogener blir kalt 'prosesserte pseudogener, fordi de mangler introner', ikke-protein kodende sekvenser. For slike gener har evolusjonister hevdet at prosesserte pseudogener oppsto fra mRNA-sekvenser som var kopiert fra et gen, ble reversert transkripert (skrevet av) i en DNA-kopi, og så gjeninnsatt i genomet. Et annet navn som ofte er brukt for denne type av pseudogener, er 'retrogener'. Til tross for dette har vitenskapsfolk identifisert viktige funksjoner for disse sekvensene i pattedyr siden 1985 (6).
Bilde 4. Såkalt prossesert pseudogen eller 'retrogen'
Nylig ble det gjort en viktig oppdagelse om en livs-oppholdende funksjon for et menneskelig 'retrogen': PPM1K (7). Da kreftpasienters celle ble undersøkt for nivå av PPM1K, ble det funnet å være urimelig lavt, sammenlignet med friske menneskers. RNA knyttet til PPM1K ble funnet ikke bare å regulere en protein-kodende versjon av PPM1K-genet, som er involvert i å motvirke kreftsvulster, men også i et annet gen (NEK8), som ofte er assosiert med kreftcellers vekst. Åpenbart, om PPM1K pseudogenet ikke blir skikkelig regulert, men splittet/oppløst av en mutasjon, så fører det til dysfunksjon i cellesyklusen, og kreft blir resultatet.
Et annet velkjent menneskelig pseudogen, er PTENpg-pseudogenet, som fungerer, lest både forfra og bakfra (sml. palindrom), som del av et høyst komplekst gen-regulerende nettverk, som hjelper å kontrollere ogregulere cellevekst i kroppen (8). PTENpg koder for minst to ulike varianter av regulerende RNA-transkript, som del av  det komplekst regulerte genet. PTENpg koder også for det samme på motsatt side av genet, men da lest i motsatt rekkefølge! Det nevnte RNA-transkript regulerer en klasse av små regulerende molekyler, microRNAs, spesielt microRNAs som regulerer det PTEN protein-kodende genet. Dette forbløffende genet koder RNA-avskrifter (transcripts) i to ulike retninger, på en dynamisk og koordinert måte. Når mutasjoner forekommer i pseudogenet, vil ikke virkemåten fungere skikkelig. Resultatet vil være dysfunksjon og/eller kreft i cellen. Det er vanskelig å forestille seg hvordan et komplisert og livsoppholdende gen, kunne ha utviklet seg fra et uhell i evolusjonen. Og det er visselig intet genomisk fossil.
det komplekst regulerte genet. PTENpg koder også for det samme på motsatt side av genet, men da lest i motsatt rekkefølge! Det nevnte RNA-transkript regulerer en klasse av små regulerende molekyler, microRNAs, spesielt microRNAs som regulerer det PTEN protein-kodende genet. Dette forbløffende genet koder RNA-avskrifter (transcripts) i to ulike retninger, på en dynamisk og koordinert måte. Når mutasjoner forekommer i pseudogenet, vil ikke virkemåten fungere skikkelig. Resultatet vil være dysfunksjon og/eller kreft i cellen. Det er vanskelig å forestille seg hvordan et komplisert og livsoppholdende gen, kunne ha utviklet seg fra et uhell i evolusjonen. Og det er visselig intet genomisk fossil.
Bilde 5. Eks. på palindrom-under BINGO
Antallet mulige pseudogener har i løpet av få år, sunket fra 95% til under 20%, og synker fortsatt. Så hvorfor feiler evolusjonære forutsigelser så ofte i forbindelse med pseudogener? Grunnen er at evolusjonære forutsigelser ser genomet som et produkt av tilfeldig prøvende og feilende prosesser. Det mest produktive synet på et genom fra et forskningsperspektiv, ville være å forvente vedvarende funksjonalitet og utrolig bio-engineering, som produkt fra en allmektig og vis Skaper.
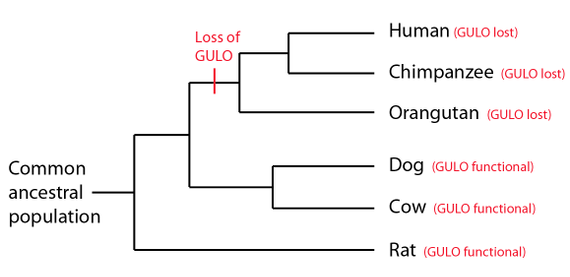 GULO pseudogenet og genetisk entropi.
GULO pseudogenet og genetisk entropi.
{Kort om entropi: Økende entropi i et system, innebærer samtidig at man taper informasjon om systemet. Den franske fysiker Léon Brillouin tolket entropi som et mål for manglende informasjon om et system. Termodynamikkens 2. hovedsetning kan da oppfattes som et uttrykk for at alle fysiske prosesser i et isolert system, fører til tap av informasjon om systemet.}
Bilde 6. GULO pseudogen
Evolusjonister har promotert GULO-pseudogenet som bevis for delte feil mellom sjimpanse og menneske, i det de har benyttet et lite antall av høyst pre-selekterte data. Deres bevisførsel er knyttet til enkelte innsettinger i ett ekson, knyttet til et fungerende GULO-gen i rotte, ikke i relasjon til en hel sekvens av GULO-gener. Vi skal se under at dette ikke 'holder vann'. I motsetning til evolusjonære påstander om ødeleggelser av vitamin C -og dets nettverk, så er de artsmessig begrenset i sine genomiske signaturer (10). De viser kjennetegn på å være uavhengige begivenheter. I en nyere studie av forfatteren til dette stoffet, så viser genomiske data at multiple tap av GULO-pseudogener i menneske, sjimpanse, og gorilla opptrådte uavhengig i hver art. Disse er assosiert med degenererte regioner, som inneholder en vid variasjon av ombyttbare gjentagende sekvenser (10). Ved reduksjonsdeling (meiosis) kan det skje uforutsette ting, som f.eks. å gjøre om sekvens-sletting i gener. GULO gen-regionen synes å ha blitt påvirket av dette. Kombinert med det faktum at mutasjoner i dette (vitamin-C) genet, ikke ville være dødelig, vil et mutert gen bli tolerert og sendt videre til etterkommerne. I denne sammenheng er det et poeng at ombyttbare elementer i det området der GULO-gen eksoner gikk tapt, er unike for mennesker, sjimpanse og gorilla, noe som indikerer at begivenhetene skjedde uavhengig av hverandre.
Dette voldte at forfatterne i et nylig evolusjonær forskningsprosjekt, valgte å satse på ID; -nei, tuller :-) I stedet oppga de fullstendig å bruke de store segmenter i GULO-gen regionen til artsmessig forskning (11). De benyttet i stedet segmenter som ga data passende for standard slutninger til evolusjonsmodellen. Altså det samme oppsettet med pre-selekterte data, brukt i så mange andre studier (cherry-picking).
Sammenlignet med sjimpanse og gorilla, er de 28.800 basene i menneskelig GULO pseudogen-region bare h.h.vis. 84% og 87% like (10). Og de 13.000 basene med DNA som går forut for levninger av menneskelig GULO-gen, -knyttet til tap av to hoved-eksoner, er bare 67% og 73% like h.h.vis. sjimpanse og gorilla. Disse resultatene ligger utenfor intervallet til felles avstamnings paradigmet og gorillaen er betydelig mer lik menneske i denne regionen, enn sjimpanse er, noe som ytterligere motbeviser den vanlig oppfattede artsmessig avstamningen.
På grunn av små endringer i ekson-regionen til menneskelig GULO-gen, det er få eller ingen, så har begivenheten antagelig nylig funnet sted. Om det skjedde for millioner av år siden, ville eksonene vært mye mer nedbrutt. I tillegg indikerer nåværende ENCODE-data i menneskelig GULO-region at det epigenetiske maskineriet til genomet har gjenkjent og tagget det som ødelagt og har lukket det ned fra å bli transkribert, for å beskytte tilhørende regioner som inneholder aktive gener (10).
GULO pseudogenet er er klart kjennetegn på genetisk diskontinuitet mellom mennesker og aper, ikke på arv av felles mistak. Artsmessig bestemte gen-degraderings-begivenheter dukker opp som et felles tema, assosiert med genetisk entropi og diskontinuitet mellom artene, ikke med makro-evolusjon.
 Oppsummering: Pseudogener svikter det evolusjonære paradigmet
Oppsummering: Pseudogener svikter det evolusjonære paradigmet
En av de evolusjonære 'sannheter' som havner i grøfta under oppvåkning av ny vitenskap, er forestillingen om at pseudogener bare er ødelagte gener. Da vitenskapsfolk fant gener som syntes å ha kodings-feil, som forhindret dem fra å lage fungerende proteiner, så ble de karakterisert som de funksjonelle levninger, eller genom-fossiler. Men nylig har forskere funnet at mange av disse såkalte fossile sekvensene, ikke er skinn/uekte (pseudo) -gener når det kommer til stykket. De er nødvendige for å opprettholde sunne livsprosesser i cellen. Og i de få tilfellene hvor reelle ødelagte gener er oppdaget, så er de ofte forbundet med arvelige sykdommer. I motsetning til vanlig evolusjonær oppfatning at delte feil og pseudo-gener stammer fra et felles opphav, så er aktuelle genom-bevis for beta-globin genet, 'retro-gener' og GULO-pseudogenet i motstrid med dette.
Bilde 7. Aktuell bok om emnet
Oversatt og omsatt til .htm-format ved Asbjørn E. Lund